上海建平实验中学(地杰校区) 200120
胃癌指源于胃黏膜上皮畑胞的恶性肿瘤,主要是胃腺癌。它是目前全世界范围内发病率最高的癌症之一,死亡率位居第三。世界卫生组织统计显示,2018 年全球胃癌的发病例数已超 103 万,其中,我国为胃癌高发国家,占全球胃癌发病例数的44%;总死亡人数逾78 万,约一半为中国人[1]。胃癌早期发现较为困难,许多患者易把早期胃癌当为胃病。据统计,百分之九十罹患胃癌的患者发现是胃癌时就已经是中晚期,使胃癌治愈率低。如何提高胃癌治愈率始终是医学领域的重点问题。已有大量文献研究了胃癌的治疗方法,但仍需要更多的研究来弥补空白。
c-Myc 是一种核内磷酸化蛋白,结构上包含非特异性DNA 结合区、碱性区、核内 DNA 结合区及转录激活区[2],可直接结合基因启动子区的E-box调控多种基因的表达,主要是通过调节多达15%的人类基因从而参与细胞周期以及细胞生长、凋亡、分化和代谢等关键进程,其表达水平受到生长因子依赖信号的严格控制,所以在正常细胞中一般很少表达或不表达,而在肿瘤细胞中则会通过多种机制来解除调控并增强其活性[3]。所以c-Myc同时也是一种致癌转录因子,在促进肿瘤发生、维持肿瘤细胞生长增殖与分化、血管形成和凋亡中发挥着重要作用。许多研究者在恶性肿瘤中都发现了c-Myc的异常表达。c-Myc的扩增对肿瘤的发生发展和转移密切相关[4-5]。其异常表达主要通过基因扩增、染色体转位、单核苷酸多态性或c-Myc相关通路的突变等机制实现的[6]。所以,许多研究者试图研发c-Myc的抑制剂作为抗癌制剂,c-Myc在肿瘤中的作用和研究受到越来越多的关注。
同时,一些信号通路也与肿瘤的发生有着许多关联。其中,与胃癌相关的有TGF/β通路、Hippo/YAP通路、Wnt通路、PI3K/AKT通路、Ras/MAPK通路等。信号通路是由于一些在细胞外产生的信号通过细胞膜传入细胞内部从而发挥生物学效应的一种酶促反应通路。信号通路的蛋白组成不同,其发挥的生物学效应也不同。目前研究证明,Ras/MAPK信号通路在肿瘤细胞的增殖、凋亡、血管形成等过程中均发挥着至关重要的作用。该通路的异常激活会促进肿瘤细胞异常增殖分化,抑制细胞凋亡,增强细胞对缺氧和营养不良等环境的耐受性,辅助肿瘤细胞生长,促进肿瘤细胞转移[7]。
前人已有研究发现,胃特异性c-Myc转基因(Atp4b-Cre; Myc OE/+)小鼠可以产生胃癌表型,并伴随着PI3K-AKT途径激活[8]。所以,在本研究中,我们聚焦于癌基因c-Myc驱动的小鼠胃癌模型中的Ras/MAPK途径分子信号检测,在分子水平上研究肿瘤发生前后小鼠胃癌模型中Ras/MAPK信号通路中的因子变化,旨在探索Ras/MAPK信号通路在胃癌发生发展过程中的作用,为胃癌的进一步诊断、治疗的基础科学研究提供思路。
材料
氢氧化钠、EDTA、二甲苯Ⅰ、 二甲苯Ⅱ、无水乙醇Ⅰ、无水乙醇Ⅱ、95%乙醇Ⅰ、95%乙醇Ⅱ、90%乙醇、80%乙醇 、70%乙醇 、苏木素 、伊红、中性树胶、GelRed、1X TAE、琼脂糖、5X PrimeScript Buffer、PrimeScript RT Enzyme Mix Ⅰ、Oligo dT Primer、Random 6 mers、RNA、RNase Free dH2O 、SYBR Green、Primer mix、ddH2O、cDNA
鼠尾DNA提取
鼠尾裂解液A:称取0.1g NaOH 粉末和0.006 g EDTA 粉末,溶于100mL ddH2O中。
鼠尾裂解液B:称取0.484g Tris 粉末,溶于50ml ddH2O 中,调节pH 至5.5,最后用ddH2O 补足至100mL。
剪取胃特异性转基因c-Myc的小鼠鼠尾,约2 mm 鼠尾放入1.5 ml 离心管中,加入100 μL 鼠尾裂解液A,98℃金属浴1 h,短暂离心以使管盖上液体流下,加入同等体积的鼠尾裂解液B 中,得胃特异性转基因c-Myc的小鼠DNA样本。
基因型鉴定
配制15 μL的PCR体系,包含Premix Taq 酶 7.5 μL、正向引物 0.5 μL、反向引物 0.5 μL、ddH2O 6 μL、DNA 0.5 μL。进行PCR鉴定小鼠基因型,其中,PCR 反应程序预变性为94℃ 2 min。PCR变性94℃ 20 s,退火58℃ 30 s,延伸72℃ 20 s,进行35个循环,并延伸72℃ 2 min,保持10℃ 。将琼脂糖溶于1X TAE 中配置2%的DNA 琼脂糖凝胶,依据配置体积在微波炉中适当加热几min至溶液完全澄清。待稍微冷却后,加入GelRed核酸染料(1:10000),混匀后倒胶。室温下冷却凝固后,将凝胶放入电泳槽中,点样,8 μL /孔。打开电泳仪,接好电极,120 V恒压跑胶40 min。于凝胶成像仪中照胶,拍照记录,依据条带大小鉴定基因型。
表1 小鼠基因型鉴定所用的PCR引物序列
引物名称 | 引物序列 |
cre-F | GCCTGCATTACCGGTCGATGCAACGA |
cre-R | GTGGCAGATGGCGCGGCAACACCATT |
c-Myc-wt-F | CCAAAGTCGCTCTGAGTTGTTATC |
c-Myc-wt-R | GAGCGGGAGAAATGGATATG |
c-Myc-mut-F | CCAAGAGGGTCAAGTTGGA |
c-Myc-mut-R | GCAATATGGTGGAAAATAAC |
实时荧光定量PCR 法(Q-RT-PCR法)
配置10 μL的反转录体系,包含2 μL 5X PrimeScript Buffer (for Real Time)、0.5 μL PrimeScript RT Enzyme Mix Ⅰ 、0.5 μL Oligo dT Primer 、0.5 μL Random 6 mers、500ng RNA 、10 μL RNase Free dH2O。进行37℃水浴15 min, 85℃水浴10 s,从水浴锅中拿出离心管,加入90 μL 的ddH2O 至体系中得到cDNA,将cDNA 样品保存在-20℃冰箱中。配置10 μL的qPCR 体系,包含SYBR Green 5 μL、Primer mix 0.4 μL、ddH2O 3.6 μL、cDNA 1 μL 每个实验孔保证至少有3 个以上重复,以GAPDH 作为内参对照,加样完贴膜,随后3000rpm 离心2 min。放入实时荧光定量PCR 仪中进行PCR,其中,PCR 程序变性为95℃ 30 s,PCR进行95℃ 5 s,60℃ 30 s,40个循环后,95℃ 15 s,融解60℃ 15 s,95℃ 15 s。PCR 程序结束后得到各个孔的Ct 值,根据标准曲线得到各孔的初始模板量,由此判断各个基因的RNA 表达水平。
表2 Q-RT-PCR法所用的PCR引物序列
引物名称 | Forward Primer | Reverse Primer |
rasgrf2 | CAGAAGAGCGTGCGCTACAA | TCTGGTAGAGAGCGAACCACT |
akt1 | ATGAACGACGTAGCCATTGTG | TTGTAGCCAATAAAGGTGCCAT |
akt2 | ACGTGGTGAATACATCAAGACC | GCTACAGAGAAATTGTTCAGGGG |
β-actin | GGCTGTATTCCCCTCCATCG | CCAGTTGGTAACAATGCCATGT |
c-Myc | GAACAAGAAGATGAGGAAGAA | CAGAAGGTGATCCAGACT |
fgf14 | GGCTTGATCCGTCAGAAACG | CACCAGGTTGCCATTGAAAAG |
组织学染色
事先准备好盖玻片。脱蜡:二甲苯Ⅰ、Ⅱ各10min。复水:100%(Ⅰ、Ⅱ)、90%、80%、70%乙醇各5min,自来水冲洗3遍,每次5min。苏木素染色8min,流水冲洗10min。 伊红染色1min,流水冲洗10min。脱水:70%、80%乙醇各30秒,90%乙醇1min、95%乙醇 、100%乙醇Ⅰ、100%乙醇Ⅱ 各2min。滴上中性树胶,封片。正置显微镜下镜检,并进行图像采集。
统计学分析
本研究主要运用EXCEL作图法,结合t检验,对所得结果进行分析。
c-Myc胃特异性转基因小鼠的基因型鉴定
本研究采用DNA凝胶电泳法对所研究的小鼠基因型进行鉴定,结果如下图所示,第一条泳道为marker,第二三四条泳道为Atp4b-Cre; Myc OE/+,第五六七条泳道为Myc OE/+,最后三条泳道为WT小鼠。结果表明成功鉴定出Atp4b-Cre; 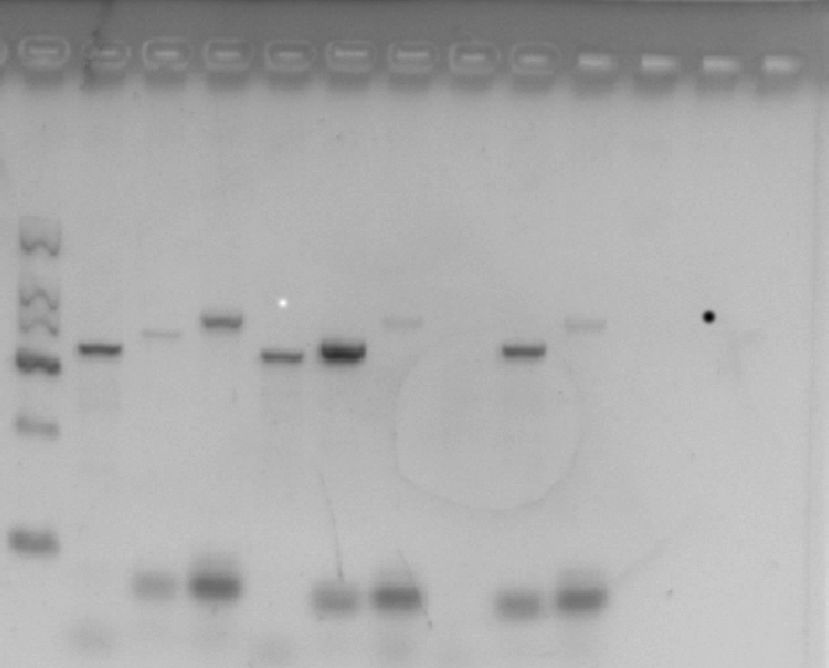 Myc OE/+小鼠。
Myc OE/+小鼠。
图1 DNA凝胶电泳鉴定结果
RAS/MAPK分子信号通路检测
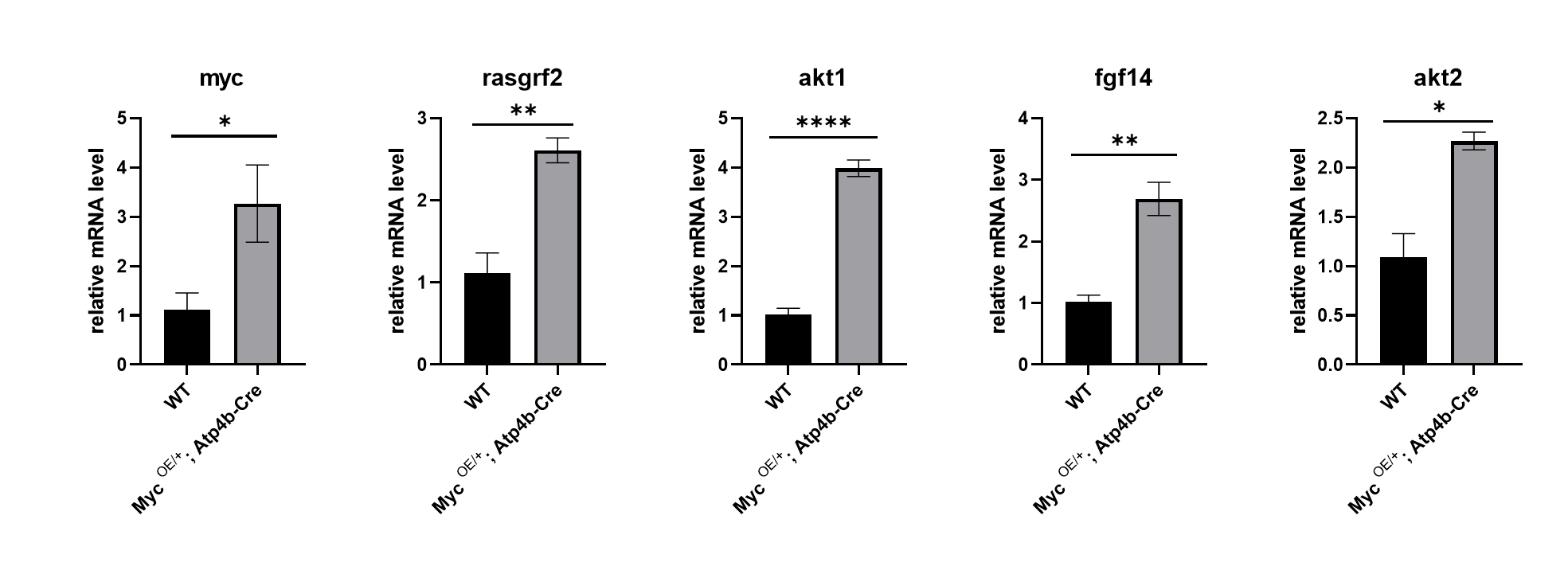 本研究利用rasgrf2,akt1,akt2,β-actin,c-Myc,fgf1引物序列,采用Q-RT-PCR法对正常组小鼠和c-Myc转基因小鼠进行了相关信号分子通路的检测,qPCR结果如图2所示,结果表明,与正常组小鼠相比,RAS/MAPK通路中相关基因都增加了2-3倍,具有显著性差异,这些结果说明RAS/MAPK通路出现了上调现象。
本研究利用rasgrf2,akt1,akt2,β-actin,c-Myc,fgf1引物序列,采用Q-RT-PCR法对正常组小鼠和c-Myc转基因小鼠进行了相关信号分子通路的检测,qPCR结果如图2所示,结果表明,与正常组小鼠相比,RAS/MAPK通路中相关基因都增加了2-3倍,具有显著性差异,这些结果说明RAS/MAPK通路出现了上调现象。
图2 RAS/MAPK信号转导相关基因的Q-RT-PCR结果
组织染色
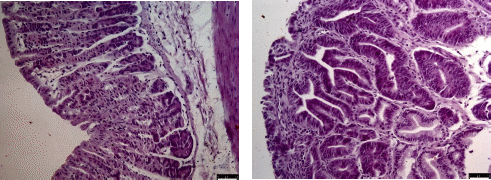
本研究采用HE染色法观察了正常组小鼠与C-Myc过表达小鼠组的胃切片,结果如下图所示,正常组小鼠胃黏膜完整、腺体结构正常,细胞排列整齐,与正常组小鼠相比,c-Myc过表达小鼠胃体腺体结构出现异常增生,细胞核质比增大,细胞排列混乱,癌细胞向黏膜下层浸润。综上,c-Myc过表达小鼠出现了严重的胃癌表型。
图3 组织染色结果(WT左,Atp4b-Cre; MycOE右)
结论:本研究通过DNA凝胶电泳技术成功鉴定出Atp4b-Cre; Myc OE/+小鼠,利用Q-RT-PCR法检测了RAS/MAPK通路中相关基因的表达情况,最后采用HE组织染色技术观察了正常小鼠与Atp4b-Cre; Myc OE/+小鼠的胃组织切片,研究结果表明C-Myc驱动的胃癌中,RAS/MAPK信号通路发生了上调。
rasgrf2、akt1、akt2、fgf14 都是RAS/MAPK信号通路中的重要成员,本研究中检测到这些相关基因的表达上调都提示RAS/MAPK信号通路发生了上调。Rasgrf2是细胞膜信号传导通路中的重要分子,能够影响细胞的生长、分化、转移及凋亡,能使与GDP结合而失活的Ras蛋白释放GDP,恢复Ras至活性状态,在Ras信号途径中起着重要的调控作用,参与体内多种生理功能。fgf14是成纤维细胞同源性生长因子,在血管生成中发挥重要的调节作用,fgf诱导体内新血管的产生,在胚胎期和伤口愈合过程中新血管的发育中发挥重要作用。在体外,fgf通过与其特异性的受体结合,参与细胞增殖、迁移以及蛋白酶的产生。akt1是一种丝氨酸蛋白酶,广泛分布在组织中,参与多种生物学过程,包括代谢,增殖,细胞存活,生长,胰岛素信号传导和血管生成等。akt1基因的突变也被证明在体外赋予对变构激酶抑制剂的抗性。akt2是一个假定的癌基因,主要分布在肌肉和脂肪组织中,许多恶性肿瘤的发生发展与akt2的扩增、过表达和激酶活性升高有关,akt2主要介导肿瘤细胞的粘附、运动、侵袭和转移效应,其表达水平与肿瘤的发生、发展、预后及化疗敏感性密切相关。对PI3K/Akt信号通路的激活起到重要作用。
在本研究中,RAS/MAPK分子信号通路发生了上调,Ras 蛋白是调节细胞生长和增殖信号通路的重要元件,提示胃癌的发生机制可能为:当Ras变异时,将信号转导到下游信号元件可能引起细胞的异常增殖,导致胃癌的发生。生长因子受体介导的Ras/MAPK信号转导途径是诸多信号途径中与细胞增殖、分化密切相关的重要信号途径。多肽生长因子与受体结合,激活下游信号分子如Ras、丝裂原激活蛋白激酶( MAPK)等,通过级联酶促反应,影响基因表达,调控细胞增殖和分化。本研究再次从动物模型的层面来分析胃癌中相关信号通路的变化,RAS/MAPK信号通路发生上调,从而促进了胃癌细胞的增殖。本研究为进一步研究胃癌中其他分子信号提供了基础,为胃癌的诊断与治疗等提供了一定的理论依据。
在本次研究中,由于对于实验步骤及要点的不熟悉,出现了一些操作上的失误,例如忘记添加核酸染料,添加了错误的试剂,仪器规格错误等问题。 进行DNA凝胶电泳鉴定时,在将DNA注射进入琼脂糖凝胶中需要小心,此操作不当容易造成刚注射进去的DNA被吸上来,最后造成实验误差。在对切片进行染色时,由于操作失误造成切片染色不完全,对实验结果造成一定影响。在对组织切片进行染色时,染色时间需要在一定范围,时间过长容易导致染色过深,时间过短会导致不上色。这些问题在今后的工作中务必引起重视。
1 参考文献
[]Serra O, Galán M, Ginesta MM, et al. Comparison and applicability of molecular classifications for gastric cancer[J]. Cancer Treat Rev, 2019, 77:29-34.
2[]邹治铭, 郭华雄. C-myc在肿瘤中作用的研究进展[J]. 现代医学, 2016(7):3.
3[]刘雅惠, 高露, 王亚菁,等. c-Myc调控肿瘤代谢作用机制的研究进展[J]. 中国药科大学学报, 2021, 52(3):8.
4[]Xia S, Ma J, Bai X, et al. Prostaglandin E2 promotes the cell growth and invasive ability of hepatocellular carcinoma cells by upregulating c-Myc expression via EP4 receptor and the PKA signaling pathway[J]. Oncol Rep, 2014, 32(4):1521-1530.
5[]Nair R, Roden DL, Teo WS, et al. c-Myc and Her2 cooperate to drive a stem-like phenotype with poor prognosis inbreast cancer[J]. Oncogene, 2014, 33(30):3992-4002.
6[]Lin, Charles Y.; Lovén, Jakob; Rahl, Peter B.; Paranal, Ronald M.; Burge, Christopher B.; Bradner, James E.; Lee, Tong Ihn; Young, Richard A. (2012). Transcriptional Amplification in Tumor Cells with Elevated c-Myc. Cell, 151(1), 56–67.
7[]袁莉, 贾莉. PI3K/AKT/PTEN通路与肺癌的关系[J]. 齐齐哈尔医学院学报, 2013, 34(18):2657-2659.
8[] Liu Jing et al. Stomach-specific c-Myc overexpression drives gastric adenoma in mice via AKT/mTOR signaling.[J]. Bosnian journal of basic medical sciences, 2020.

客服QQ:30444492琼网文【2021】1550-113号
增值电信业务经营许可证:琼B2-20210322
出版物经营许可证:新出发龙华出字第(2021)009号
广播电视节目制作经营许可证:(琼)字第00779号
版权所有 ©2002-2024 期刊网(www.qikanchina.com) 琼ICP备2021005105号



