广西柳州市工人医院 545005
【摘要】目的:探讨siRNA 沉默HnRNP L基因对RWPE-1细胞、Lncap细胞、PC3细胞生物学行为的影响。方法:通过合成了靶向HnRNP L的特异性siRNA,并通过了Western Blot在蛋白水平上进行检测,并筛选出干扰效率最高的3号序列来干扰RWPE-1细胞、Lncap细胞、PC3 细胞中HnRNP L的表达,观察其对三种不同细胞生长、凋亡、体外侵袭能力及迁移能力的差异。结果:沉默HNRNPL后,CCK-8实验提示三种细胞的增值都明显降低,划痕实验结果提示:沉默HNRNPL后,RWPE-1细胞和PC3细胞的迁移力明显减弱。Transwell检测结果提示:沉默HNRNPL后,RWPE-1细胞、Lncap细胞和PC3细胞的侵袭能力明显减弱。结论:HnRNPL对前列腺癌的多种生物学行为有明显的影响,参与了前列腺癌的发生、发展的恶性进程,促进了前列腺癌的生长、侵袭和转移。
【关键词】HnRNPL蛋白; siRNA; CCK-8、划痕实验、Transwell检测
【Abstract】Objective: To investigate the effect of siRNA silencing HnRNP L gene on the biological behavior of RWPE-1 cells, Lncap cells and PC3 cells. Methods: The specific siRNA targeting HnRNP L was synthesized and detected at the protein level by Western Blot, and the No. 3 sequence with the highest interference efficiency was screened to interfere with HnRNP in RWPE-1 cells, Lncap cells and PC3 cells. The expression of L was observed, and its differences in the growth, apoptosis, in vitro invasive ability and migration ability of three different cells were observed. Results: After silencing HNRNPL, the CCK-8 experiment showed that the proliferation of the three cells was significantly reduced, and the scratch experiment results showed that: after silencing HNRNPL, the migration ability of RWPE-1 cells and PC3 cells was significantly weakened. The results of Transwell assay showed that the invasive ability of RWPE-1 cells, Lncap cells and PC3 cells was significantly weakened after silencing HNRNPL. Conclusion: HnRNPL has obvious effects on various biological behaviors of prostate cancer, participates in the malignant process of occurrence and development of prostate cancer, and promotes the growth, invasion and metastasis of prostate cancer.
【Key words】HnRNPL protein; siRNA; CCK-8, scratch assay, Transwell assay
前列腺癌目前是男性常见的恶性肿瘤,居西方国家男性所有恶性肿瘤发病率的第二位,死亡率的首位[1]。而在美国,前列腺癌的发病率则位居男性所有恶性肿瘤的首位,死亡率仅次于肺癌[2]。我国前列腺癌的发病率虽比西方国家低很多,但随着人口的老龄化、生活水平的提高以及医疗的改善呈现明显上升的趋势
[3],对我国男性的健康已经有很大的影响。
核不均一蛋白L(hnRNP L)是由Piñol-Roma S[3]等在早期转录片段中发现的富含胱氨酸及脯氨酸的核糖核蛋白,是核不均一核糖核蛋白(核蛋白)复合体的一个重要组成部分[4]。我们前期实验已经证实了HnRNP L蛋白在前列腺癌中是高表达,与前列腺癌病理分级是正相关,与临床分期无关。同时HnRNP L蛋白在PC3细胞中表达最高,在RWPE-1细胞中表达最低。同时hnRNPs家族中的HnRNP K已被报道跟前列腺癌有着密切关系,并参与前列腺癌的增值、分化及凋亡过程[5]。
同时,当将某特定基因mRNA 同源的siRNA 导入到细胞内时,而该同源基因的mRNA 就可能发生降解或者蛋白质翻译被抑制;从而出现基因沉默现象,最终导致该基因的一些功能发生改变,进而该细胞的生物学行为也开始发生变化。因此我们拟通过合成了靶向HNRNPL的特异性siRNA,并通过了Western Blot在蛋白水平上进行检测,并筛选出干扰效率最高的序列来干扰RWPE-1细胞、Lncap细胞、PC3 细胞HnRNP L的表达,观察其是否引起生物学行为的变化,结合文献报道,我们重点观察了细胞生长、凋亡、体外侵袭能力及迁移能力的变化。
材料与方法
材料. 正常前列腺上皮细胞(RWPE-1)由中山大学附属第三医院高新教授馈赠;激素依赖性前列腺癌细胞(Lncap)及激素非依赖性前列腺癌细胞(PC3)由南方医科大学南方医院赵善超教授馈赠。CCK-8购自于日本DOJINDO公司;Matrigel胶购自于美国BD公司;Transwell Chamber(8μm孔径,3422)购自于美国Corning公司;荧光显微镜Q550CW购自于德国Leica公司.
细胞培养 先从液氮罐里取出细胞冻存管,然后进行将细胞快速转入到37℃~38℃的水浴中解冻复苏,将细胞放入培养瓶中,把培养瓶放在5%C02、37℃恒温培养箱中培养,一般等24-48h后待细胞很大部分都贴壁就可以开始换液,必要时进行传代。
1.3实验分组
实验分A、B、C三大组,每大组中分三小组:
A组:
第一组(NC): 阴性对照组:RWPE-1细胞+NC-siRNA
第二组(3号siRNA):干扰组:RWPE-1细胞+ HNRNPL –siRNA3
第三组(F):荧光标记组:RWPE-1 细胞+FAM-NC-siRNA
B组:
第一组(NC): 阴性对照组:Lncap 细胞+NC-siRNA
第二组(3 号siRNA):干扰组:Lncap 细胞+ HNRNPL –siRNA3
第三组(F):荧光标记组:Lncap 细胞+FAM-NC-siRNA
C组:
第一组(NC): 阴性对照组:PC3细胞+NC-siRNA
第二组(3 号siRNA):干扰组:PC3细胞+ HNRNPL –siRNA3
第三组(F):荧光标记组:PC3 细胞+FAM-NC-siRNA
将三种细胞分别加入6孔培养板,按实验分组要求加入转染复合物;加入转染复合物6小时后去掉培养基,换成1640完全培养基,2ml/孔,继续温育;转染6小时以后先观察F组FAM标记的阴性对照序列的荧光图片,荧光显微镜下观察如果转染效率在80%以上时,可以进行其他后续实验。
1.4 CCK-8法实验步骤:
(1). 选生长状态良好的对数生长期细胞,先用胰酶细胞后消化离心,调整细胞的浓度并根据实验分组要求分组,将重悬好的单细胞接种于6孔板中,在37℃、5% CO2 恒温培养箱中温育24小时后开始转染细胞,细胞培养和转染方法同第二部分;
(2). 转染后将各组细胞经胰酶消化后离心,重悬单细胞调至终浓度为2×105个/ml后接种于96孔板,根据实验分组要求分为2组,每组设4个时间点,分别为:0h、24 h 、48 h 、72 h,每个时间点设立5复孔,每孔体积为100μl;
(3). 将各组细胞放入37℃、5% CO2 恒温培养箱中继续培养。
(4). 每个时间点取出一块相应的96 孔板,每孔加入CCK-8 液10μl,放入培养箱中继续培养0.5-4 小时。
(5). 然后取出96孔培养板,放在酶联免疫检测仪上测定其光吸收值,取450nm 波长,计算每组5个复孔的平均值。
1.5各组细胞迁移力的检测(划痕实验)
(1). 选生长状态良好的对数生长期细胞,先用胰酶细胞后消化离心,调整细胞的浓度并根据实验分组要求分组,将重悬好的单细胞接种于24孔板中,在37℃、5% CO2 恒温培养箱中温育24小时后开始转染细胞,细胞培养和转染方法同第二部分,同时每组样本设2 个复孔;
(2). 转染完成后将24孔板放到37℃、5% CO2 培养箱中继续培养;
(3). 24h 后取出24孔板,吸走培养基,用10ul经过高压过的枪头在24孔板底部划痕,划痕后用PBS液洗涤细胞三次,加入含5%FBS 的1640 培养基500ul;
(4). 将24孔板放到37℃、5% CO2 恒温培养箱中培养,并于划痕后0h、24h、48h 于100 倍显微镜下拍照;
(5). 统计同一视野下裂痕区上的细胞数,同时计算每组2个复孔的平均值。
1.6 Transwell侵袭实验:其实原理简单地说就是用一层膜将高营养的培养液和低营养的培养液隔开,细胞放在低营养的培养液里,为了找吃的,细胞会往高营养的培养液里面跑,但是有膜挡着,所以要穿过膜才行。我们在膜上涂上一层基质胶,模仿细胞外基质,于是细胞要把基质消化了才可以从低营养的培养液跑到高营养的培养液里面,最后我们检测高营养的培养液里细胞量就可以知道细胞的侵袭能力了。
(1). 细胞培养和转染方法同第二部分;
(2). 转染完成后将装有各组细胞的6孔板放到37℃、5% CO2培养箱中培养48小时;
(3). 在实验的前一天晚上将Matrigel从-20℃冰箱转移至4℃冰箱,使其能够完全溶解;
(4). 为了防止Matrigel在室温下容易凝固,把与Matrigel有可能接触的实验用品在实验前1h放入-20℃冰箱中;
(5).将10μl Fibronectin涂在Transwell小室的聚碳酸脂膜的下室面上;在Transwell小室的上室面涂Matrigel 50μl,等待风干,从而形成一个基质屏障;
(6).使用前在Transwell 小室的上下室均放入约500μl 37℃的无血清1640培养基,并在温箱内静置2h后倒掉培养基;
(7). 将(2)中的细胞在实验的前24小时更换为无血清的培养基,令细胞处于饥饿状态;然后用0.25%胰酶(RWPE-1细胞用0.05%胰酶)消化后制成单细胞悬液,终浓度调为3×105/m的无血清1640细胞悬液,用移液枪吸取200μl加到Transwell细胞培养小室上室中,每组设2个复孔;
(8).同时另外取一个24孔板,每孔加入含15%FBS的1640培养基550μl,将小室浸于此板的培养基中,保证下室的培养基能接触到上室底部,恒温箱孵育24h;
(9). 将Transwell细胞小室取出,倒掉培养液,用棉签轻轻擦掉未穿过膜的细胞(滤膜上层细胞),用4%多聚甲醛固定细胞40min,再用PBS液漂洗3次,然后Giemsa染色液染色35min,流水冲洗后取下聚碳酸酯膜,待风干后于×40倍镜下计数侵袭细胞数,每个膜计数上下左右中5个不同视野所透过的细胞数,计算其平均值。
2、统计学方法
采用SPSS 25.0 软件进行统计学处理,计量资料以![]() ±s表示,采用析因分析方法分析转染前后RWPE-1细胞、Lncap细胞及PC3的细胞增殖能力的CCK-8 吸光度值(OD);采用两独立样本的t检验方法,分析干扰后不同组间CCK-8 吸光度及划痕实验的结果及Transwell穿透的细胞数, P<0.05表示具有统计学意义。
±s表示,采用析因分析方法分析转染前后RWPE-1细胞、Lncap细胞及PC3的细胞增殖能力的CCK-8 吸光度值(OD);采用两独立样本的t检验方法,分析干扰后不同组间CCK-8 吸光度及划痕实验的结果及Transwell穿透的细胞数, P<0.05表示具有统计学意义。
3、结果
3.1CCK-8法观察干扰后RWPE-1、Lncap及PC3细胞的增殖情况
经析因方差分析,瞬时下调HnRNPL表达后,RWPE-1细胞增殖能力与对照组相比显著下降,差异有统计学意义 (表3-1. F=315.064,P=0.000)。不同的时间点对RWPE-1细胞的增殖影响显著(F=431.920, P= 0.000)。各时间点组与处理组间交互效应显著(F=21.213,P=0.000);Lncap细胞增殖能力与对照组相比显著下降,差异有统计学意义 (表3-2. F=568.718,P=0.000)。不同的时间点对Lncap细胞的增殖影响显著(F=1543.436, P= 0.000)。各时间点组与处理组间交互效应显著(F=51.226,P=0.000);PC3细胞增殖能力与对照组相比显著下降,差异有统计学意义 (表3-3. F=668.116,P=0.000)。不同的时间点对PC3细胞的增殖影响显著(F=2871.830, P= 0.000)。各时间点组与处理组间交互效应显著(F=123.353,P=0.000)。
表3-1. RWPE-1细胞中HnRNPL基因沉默对其增殖能力的影响
| RWPE-1+NC | RWPE-1+siRNA | total | t/F | P |
1day | 0.773±0.075 | 0.636±0.066 | 0.705±0.098 | 3.061 | 0.016 |
2day | 1.379±0.126 | 0.732±0.079 | 1.056±0.356 | 9.725 | 0.000 |
3day | 1.862±0.058 | 1.156±0.071 | 1.509±0.378 | 17.137 | 0.000 |
4day | 2.658±0.067 | 1.859±0.195 | 2.258±0.443 | 8.660 | 0.000 |
total | 1.668±0.712 | 1.096±0.506 | 1.382±0.675 | F =315.064* | P=0.000* |
F | F =430.745 | F =115.000 | F =431.920* | | F =21.213# |
P | P=0.000 | P=0.000 | P=0.000* | | P=0.000# |
*主效应统计量及P值 #交互效应统计量及P值
表3-2. Lncap细胞中HnRNPL基因沉默对其增殖能力的影响
| Lncap+NC | Lncap+siRNA | total | t/F | P |
1day | 0.732±0.061 | 0.676±0.066 | 0.704±0.067 | 1.393 | 0.201 |
2day | 1.245±0.065 | 0.769±0.061 | 1.007±0.257 | 11.974 | 0.000 |
3day | 1.820±0.053 | 1.183±0.027 | 1.502±0.338 | 23.990 | 0.000 |
4day | 2.623±0.048 | 2.056±0.068 | 2.339±0.304 | 15.174 | 0.000 |
total | 1.605±0.723 | 1.171±0.562 | 1.388±0.676 | F =568.718* | P=0.000* |
F | F =1011.031 | F =517.401 | F =1543.436* | F =51.226# | |
P | P=0.000 | P=0.000 | P=0.000* | | P=0.000# |
*主效应统计量及P值 #交互效应统计量及P值
表3-3. PC3细胞中HnRNPL基因沉默对其增殖能力的影响
| PC3+NC | PC3+siRNA | total | t/F | P |
1day | 0.667±0.031 | 0.682±0.035 | 0.675±0.032 | -0.763 | 0.467 |
2day | 1.093±0.043 | 0.855±0.036 | 0.974±0.131 | 9.486 | 0.000 |
3day | 1.843±0.040 | 1.196±0.015 | 1.519±0.342 | 33.949 | 0.000 |
4day | 2.488±0.026 | 2.019±0.075 | 2.254±0.253 | 13.212 | 0.000 |
total | 1.605±0.723 | 1.188±0.529 | 1.355±0.645 | F =668.166* | P=0.000* |
F | F =2565.817 | F =864.753 | F =2871.830* | F =123.353# | |
P | P=0.000 | P=0.000 | P=0.000* | | P=0.000# |
*主效应统计量及P值 #交互效应统计量及P值
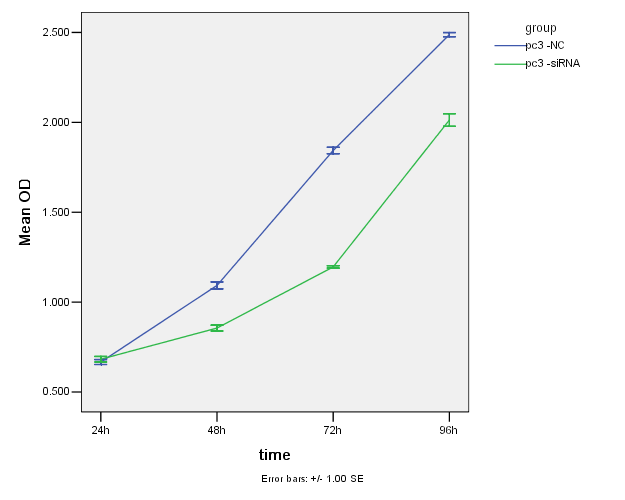
在CCK-8实验中,因用酶标仪测得的吸光值(OD值)可以间接反映活细胞数量,从下面图3-2、图3-3及图3-4可见siRNA转染24小时,RWPE-1组与对照组细胞的增值有显著性差异(P<0.05);而Lncap组与对照组无差异(P>0.05);PC3组与对照组也无差异性(P>0.05)。24至48小时三种细胞的增值都比较缓慢,而48-96小时细胞的增值速度上升;总体来说,沉默HNRNPL后,三种细胞的增值都明显降低。
图3-2. 两组RWPE-1细胞的增值曲线图
Figure 3-2. Two groups of RWPE - 1 cell growth curve
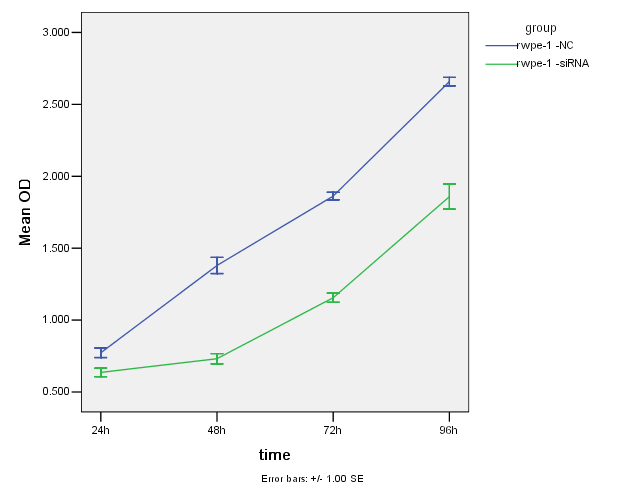
图3-3. 两组Lncap细胞的增值曲线图
Figure 3-3. Two groups of Lncap cell growth curve
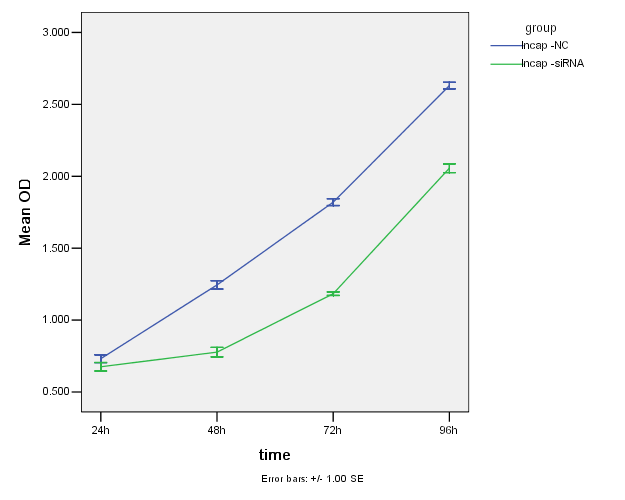
图3-4. 两组PC3细胞的增值曲线图
Figure 3-4. Two groups of PC3 cell growth curve
3.2.划痕实验检测各组细胞的迁移力
在干扰组转染后第24小时用划痕法测各组细胞的迁移力,划痕后第48小时对划痕区的细胞数进行计数(见表3-4、表3-5,图3-5、图3-6)显示:对照组迁移速度明显较快,划痕区细胞的数目明显增多,划痕区界线在划痕后第48小时已变得模糊不清,细胞迁移距离较远;而干扰组细胞有一定的迁移,但迁移速度较慢,划痕区细胞数目增加较少。比较后可得出PC3细胞干扰组的迁移力明显低于对照组,差别有统计学意义(F=4.384,P<0.01); 细胞干扰组的迁移力明显低于对照组,差别有统计学意义(F=0.808,P<0.01);实验结果提示:沉默HNRNPL后,RWPE-1细胞和PC3细胞的迁移力明显减弱。
图3-5. 划痕实验PC3细胞迁移情况(×100)
Figure 3-5. The migration of PC3 cell in Scratch experiment (×100)
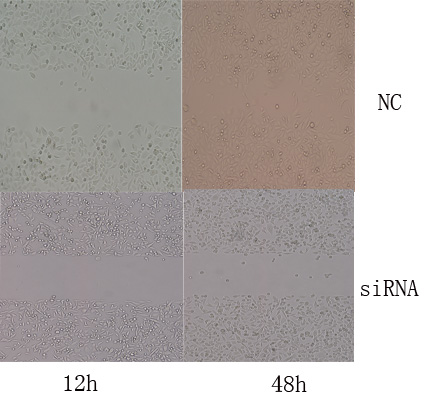
图3-6. 划痕实验RWPE - 1细胞迁移情况(×100)
Figure 3-6. The migration of RWPE - 1 cell in Scratch experiment (×100)

表3-4. 划痕后各PC3细胞组的迁移力
Table 3-4. The migration of PC3 cell after Scratch experiment

表3-5. 划痕后各RWPE - 1细胞组的迁移
Table 3-5. The migration of RWPE - 1 cell after Scratch experiment

3.3.Transwell检测各组细胞的侵袭力
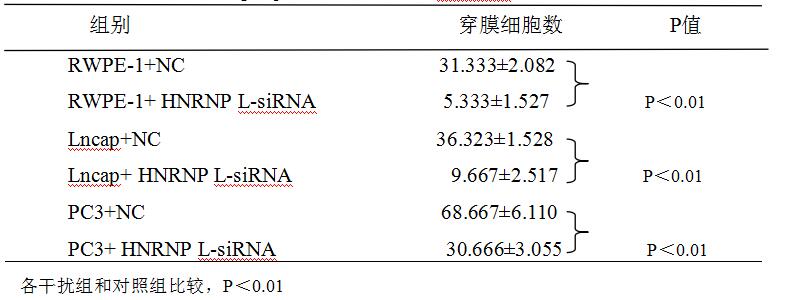
在干扰组转然后48小时用Transwell法测各组细胞的侵袭力(见如下表3-6,图3-7所示),RWPE-1细胞组干扰组穿过的平均细胞数是5个,对照组为31个,两组的差异在统计学上有显著意义(F=0.500,P<0.01);Lncap细胞干扰组平均细胞数是9个,对照组是36个,两组的差异统计学上有显著意义(F=0.643,P<0.01);PC3细胞干扰组平均穿过的细胞数是30个,而对照组是68个,两组的差异统计学上有显著意义(F=1.538,P<0.01)。从以上三种细胞的检测结果提示:沉默HNRNPL后,RWPE-1细胞、Lncap细胞和PC3细胞的侵袭能力明显减弱。
图3-7. 三种细胞穿过Transwell小室的情况
Figure 3-7. Three kinds of cells traverse the Transwell chamber
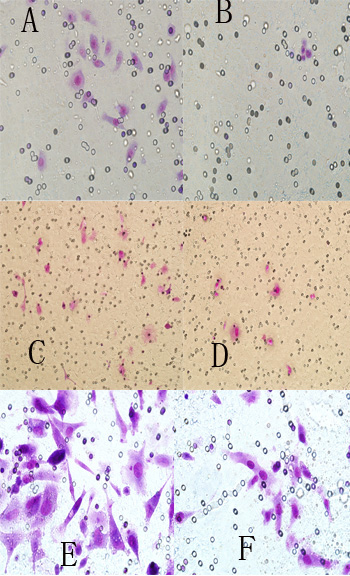
在如上图3-7中可见, RWPE-1+ HNRNP L-siRNA(B组)、Lncap+ HNRNP L-siRNA(D组)及PC3+ HNRNP L-siRNA(F组)的穿越小室能力比相应的对照组RWPE-1+NC组(A组)、Lncap+NC组(C组)及PC3+NC组(E组)明显减弱。
表3-6. 各组细胞的穿膜细胞数
Table 3-6. Each group of cells traverse the Transwell chamber
讨论
核不均一核糖核蛋白L(Heterogeneous nuclear ribonucleoprotein L , HnRNPL)是由Piñol-Roma S等在早期的转录片段中偶然发现,该蛋白富含胱氨酸和脯氨酸;是核不均一核糖核蛋白复合体的一个很重要的成员。HnRNPL在mRNA的形成、加工、包装以及维持RNA的稳定性等方面起着很重要作用。转录后加工,是调控蛋白表达的重要途径之一。HnRNPL能够与富含CA及重复CA的pre-RNA的结合,并且调控这些基因的表达。已被证实受HnRNPL调控蛋白有iNOS,bcl-2,CEACAM1 ,caspase-9及caspase-3
[11]等。
我们前期的研究证实:HnRNPL在前列腺癌中是高表达,并与前列腺癌的分化程度呈正相关,因此,HnRNPL对前列腺癌细胞的生物学行为的影响,可以帮助我们更进一步的了解HnRNPL在前列腺癌的发生、侵袭及转移等恶性进程中的作用。在干扰组转染后第24小时用划痕法测各组细胞的迁移力,划痕后第48小时对划痕区的细胞数进行计数显示:对照组迁移速度明显较快,划痕区细胞的数目明显增多,划痕区界线在划痕后第48小时已变得模糊不清,细胞迁移距离较远;而干扰组细胞有一定的迁移,但迁移速度较慢,划痕区细胞数目增加较少。比较后可得出干扰组的迁移力明显低于对照组,差别有统计学意义(P<0.01);实验结果提示:沉默HNRNPL后,RWPE-1细胞和PC3细胞的迁移力明显减弱。
运用CCK-8法测定了干扰组转染后96小时内各组PC3细胞的存活情况。CCK-8法是一种灵敏度高、重复性好的细胞活性检测方法。我们的实验结果显示:在对干扰组进行HNRNP L-siRNA转染24小时后,干扰组的细胞的增值明显慢于对照组,而且这种差异在转染后第48小时显得比较明显,然而出现这种细胞增长差异的原因主要考虑为:干扰组的细胞在lipo2000与HNRNP L-siRNA混合物结合后,24小时内就引起蛋白水平的下降,并在48小时的时候出现蛋白水平的下降变得最明显,从而引起相关的重要蛋白表达发生改变,而这些蛋白能促进细胞生长及抗凋亡等,蛋白的改变最终表现为细胞生长变慢;而第一天变化不明显主要考虑为可能转染后引起的HNRNP L-mRNA和蛋白以及生物学行为的改变存在一个效用时间延迟的问题。我们一直检测细胞转染后96小时,干扰组的细胞增长仍显著慢于对照组;因此,根据我们的实验我们可以认为沉默HNRNP L的表达,可以减少前列腺癌细胞的增值,而这种作用在细胞转染后48小时左右出现最高峰。
侵袭和转移是恶性肿瘤的两大显著特征,因此我们又进行Transwell实验来检测各组细胞的侵袭力。我们的实验中RWPE-1及Lncap细胞进入下室的细胞较少,特别是干扰组;原因主要考虑为正常上皮细胞很难像肿瘤细胞那样有很强的侵袭力,不容易穿越过小室上的聚碳酸酯膜,同时Lncap细胞相对比较难培养。从我们的实验结果可以看到干扰组细胞的侵袭力明显弱于对照组;因此,我们可以认为:沉默HNRNP L的表达,可以减少前列腺癌细胞的侵袭能力。
结合文献报道以及我们的实验结果可考虑:HnRNPL对前列腺癌的多种生物学行为有明显的影响,参与了前列腺癌的发生、发展的恶性进程,促进了前列腺癌的生长、侵袭和转移。正因为HnRNPL在肿瘤的发生、发展中起着重要的作用;因此,针对该靶点HnRNPL治疗前列腺肿瘤具有十分诱人的前景。
【参考文献】
[1] El Etrby MF,Liang Y,Lewis RW.Induction of apoptosis by mifepristone and tamoxifen inhuman LNCaP prostate cancer cells in culture.Prostate.2000;43:3-42.
[2] Jemal A,Siegel R,Xu J,et al.Cancer statistics,2010.CA Cancer J Clin,2010,60(5):277-300
[3] Pinol-Roma S,Swanson M S,Gall J G, et al. A novel heterogeneous nuclear RNP protein with a unique distribution on nascent transcripts[J]. J Cell Biol, 1989, 109(6 Pt 1):2575-87.
[4] Choi M. Identification of PABPN1 as an Interactive Protein of hnRNP L via Yeast Two-Hybrid Screening[J]. Genes & Genomics, 2008, 30(5):469-76.
[5] Suvorov A,Takser L. Delayed response in the rat frontal lobe transcriptome to perinatal exposure to the flame retardant BDE-47[J]. J Appl Toxicol, 2011, Electronic publication ahead of print.
[6] Hung L H,Heiner M,Hui J, et al. perse roles of hnRNP L in mammalian mRNA processing: a combined microarray and RNAi analysis[J]. RNA, 2008, 14(2):284-96.
[7] Matsui K,Nishizawa M,Ozaki T, et al. Natural antisense transcript stabilizes inducible nitric oxide synthase messenger RNA in rat hepatocytes[J]. Hepatology, 2008, 47(2):686-97.
[8] Lim M H,Lee D H,Jung S E, et al. Effect of Modulation of hnRNP L Levels on the Decay of bcl-2 mRNA in MCF-7 Cells[J]. Korean J Physiol Pharmacol, 2010, 14(1):15-20.
[9] Dery K J,Gaur S,Gencheva M, et al. Mechanistic control of carcinoembryonic antigen-related cell adhesion molecule-1 (CEACAM1) splice isoforms by the heterogeneous nuclear ribonuclear proteins hnRNP L, hnRNP A1, and hnRNP M[J]. J Biol Chem, 2011, 286(18):16039-51.
[10] Vu NT, Park MA, Shultz JC, et al. hnRNP U enhances caspase-9 splicing and is modulated by AKT-dependent phosphorylation of hnRNP L. J Biol Chem. 2013 Feb 8.
[11] Jingping Li, Wenbin Guo, Fei Li, et al. HnRNPL as a key factor in spermatogenesis: Lesson from functional proteomic studies of azoospermia patients with sertoli cell only syndrome. J Proteomics. 2012 Jun 6;75(10):2879-91.
基金项目:广西卫健委自筹科研项目(编号 Z20190777),课题题目:HnRNP L在前列腺癌中的功能研究

客服QQ:30444492琼网文【2021】1550-113号
增值电信业务经营许可证:琼B2-20210322
出版物经营许可证:新出发龙华出字第(2021)009号
广播电视节目制作经营许可证:(琼)字第00779号
版权所有 ©2002-2024 期刊网 琼ICP备2021005105号



